角鼻龍屬
角鼻龍屬(屬名:Ceratosaurus,發音:/ˌsɛrətoʊˈsɔːrəs/,意為「角蜥蜴」)是獸腳類恐龍的一個屬,生存年代為晚侏羅世(啟莫里階至提通階),分布範圍包含北美洲、可能還有歐洲、非洲、南美洲等地。角鼻龍是種中型、二足行走的肉食性動物,正模標本估計身長5.3或5.69公尺,凹齒角鼻龍則更大型,可達7公尺長。角鼻龍特色在於很深的頜部,其中生長著比例特長的刀片狀牙齒;鼻部中線突出一只脊狀角,眼睛上方也有一對角狀物;前肢短但功能健全,上有四指;尾部縱向很寬;沿頸部、背部至尾部中線長有一排小型突起的皮甲,並在其餘未知身體部位亦有分佈。
| 角鼻龙属 | |
|---|---|

| |
| 克里夫蘭羅伊採石場出土的角鼻龍骨架,展示於猶他州自然歷史博物館 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | 獸腳亞目 Theropoda |
| 科: | †角鼻龙科 Ceratosauridae |
| 属: | †角鼻龙属 Ceratosaurus Marsh, 1884 |
| 模式種 | |
| †鼻角角鼻龍 Ceratosaurus nasicornis Marsh, 1884
| |
| 其它種 | |
| |
| 異名 | |
| |
1884年美國古生物學家奧斯尼爾·查爾斯·馬許根據在科羅拉多花園公園屬於莫里遜組地層出土的一副近乎完整的骨骼建立了模式種鼻角角鼻龍(C. nasicornis)。花園公園的標本至今仍是該屬已知最完整的骨骼,此後僅有其他少量標本被描述。2000年根據從猶他克里夫蘭羅伊採石場與科羅拉多果城找到的兩副破碎骨骼描述了另外兩個物種:凹齒角鼻龍(C. dentisulcatus)、大角角鼻龍(C. magnicornis),但其有效性備受質疑,這三種的骨骼很可能只是同一物種的不同生長階段。1999年記載了首個幼體標本發現。2000年描述了葡萄牙洛里尼揚組的部分骨骼,成為角鼻龍在北美以外分布的新證據。坦尚尼亞、烏拉圭、瑞士亦有可能的破碎遺骸曾被提及,但目前多數學者不承認這些屬於角鼻龍。
角鼻龍成為獸腳亞目中一大分支角鼻龍類的命名依據,這個演化支在早期即與較接近鳥類的堅尾龍類分化開來,彼此互成姊妹群。在角鼻龍類中,一些學者認為與其最近緣的物種是阿根廷的銳頜龍,後者有著類似的加長牙齒形狀。英格蘭地質較古老的原角鼻龍曾一度被認為是角鼻龍的直系祖先,現在已知道兩者并无关联。
角鼻龍與異特龍、蠻龍等其他侏羅紀北美的代表性大型獸腳類共享棲地,但其數量明顯比繁盛的異特龍要罕見許多。有推測這些掠食者的食物偏好彼此有所差異,從而佔據不同棲位以減少競爭。角鼻龍可能以植食性恐龍為獵物,捕食魚類等水生獵物亦曾被提出。鼻角功能可能並非馬許最初假設的武器,而作為視覺展示物的可能性更高。
角鼻龍在19世紀末被描述為一種非常致命的代表性肉食恐龍,廣泛被當時各大古生物博物館所青睞,但到了20世紀初,暴龍的發現及媒體文化的加持,使其逐漸取代角鼻龍原本的地位,成為了現代人們對肉食恐龍的代表形象。
發現歷史
模式種與正模標本

首個標本、同時也是正模標本USNM 4735,於1883至1884年由農民費屈(Marshall Parker Felch)所發現並挖掘。[1]發現時狀態呈現關節相連、幾乎完整並含有頭骨。較顯著遺失部位包含:未知數量的脊椎、體腔除最後一節之外的所有肋骨、肱骨、雙手遠端指骨、右前肢大部分、左後肢大部分、腳部大部分。[2]:77骨骼被包覆於堅硬的砂岩之中,頭骨與脊椎在化石形成過程中嚴重扭曲變形。[2]:2,114發現地點位於科羅拉多州峽谷城的花園公園,被稱為費屈一號採石場(Felch Quarry 1),屬於莫里遜組化石蘊含量最豐富的地層遺址之一。早在角鼻龍發現以前,已在當地挖出大量恐龍化石,其中較著名的包括1883至1884年異特龍近乎完整的正模標本USNM 4734。[2]:7,114
挖掘完後,標本以船運往紐哈芬的皮博迪自然史博物館,在那裡由馬許進行研究,1884年發表了新屬新種鼻角角鼻龍(Ceratosaurus nasicornis)。[3][2]:114屬名Ceratosaurus由古希臘語的κερας/κερατος/keras/keratos(角)加上σαυρος/sauros(蜥蜴);種名nasicornis取自拉丁語的nasus(鼻子)和cornu(角);學名全意即為「鼻子長角的角蜥蜴」。[4]因標本十分完整,發表後角鼻龍很快成為全美最知名的獸腳類恐龍。1898至1899年間,該標本與其他許多由馬許描述的標本一同轉移至華盛頓特區的美國國立自然史博物館。抵達當地時,僅少部分材料有清修,隨後於1911年起一直持續到1918年底進行了其餘的清修工程。從紐哈芬到華盛頓DC的包裝及搬運過程對角鼻龍標本造成了一些程度的損傷。[2]:2,1141920年吉爾摩發表了針對來自紐哈芬的角鼻龍及其他獸腳類標本的全面性重新描述,這之中就包括了同樣採石場出土也幾乎完整的異特龍。[2]:2

馬許在他1892年的論文描繪了角鼻龍首個骨架重建圖,身長6.7公尺、身高3.7公尺。[5]吉爾摩在1920年指出,前者的重建把軀幹弄得太長,至少多加了六節脊椎。該錯誤在隨後多篇文獻中反覆提及,包括1899年(1920年才出版)邦德(Frank Bond)在奈特指導下繪製的首件生前復原圖。1901年出版更準確的復原圖是由格里森同樣於奈特監督下製作。吉爾摩於1910至1911年間裝設了正模骨架,並展示於國立自然史博館。多數早期重建都把角鼻龍描繪成尾巴拖地的直立站姿。[2]:115–116相比之下吉爾摩的重建在當時頗為前衛:[6]:276由大腿骨上端得到啟發,發現其與小腿成一定角度,而形成水平而非直立、尾部懸空、能夠奔跑的姿勢。因為化石受壓縮變形的程度嚴重,吉爾摩難以架設立體骨骼,而用淺浮雕的樣貌展示於牆面上。[2]:114部分骨骼嵌入牆面,使要進行更深入研究受到阻礙。在2014至2019年博物館恐龍展示翻修期間,館方將標本從牆面取下,[7][8]並於2019年計畫開幕的新展覽中,以一個新製作的立體複製品取而代之;原始標本則回歸博物館館藏,使學者得以徹底研究。[8]
北美其他發現
自模式種的正模發現以後,直到1960年代才再次發現另一具較重要的角鼻龍化石,是由詹姆斯·麥德森及團隊在猶他克里夫蘭羅伊採石場挖出破碎、關節脫落的部分骨骼及頭骨UMNH VP 5278標本,是體型最大的角鼻龍標本之一。[9]:211976年艾瑞克森(Thor Erikson)在科羅拉多果城發現第二個關節連接、含頭骨的標本MWC 1。[10]這是一隻相對完整的骨骼,缺乏下頜、手臂及腹肋。雖然頭骨較完整,發現時已呈關節脫落狀態,側面壓得很扁。來自一隻大型個體,但由頭骨縫線開放程度顯示,其尚未達到成熟的體型。[9]:2–3提供博物館展示的科學準確3D頭骨重建模型,製作過程很複雜,從單一件原始骨骼翻模、矯正變形部位、重建遺失部分、將骨骼模型安裝到適當位置、最後上色以還原骨骼原始色彩。[11]
果城和克里夫蘭羅伊的標本均於2000年由麥德森和塞繆爾·保羅·威爾斯在著作中進行描述,猶他標本被指定為新種凹齒角鼻龙 (C. dentisulcatus );科羅拉多標本則為大角角鼻龍(C. magnicornis)。[9]前者的種名dentisulcatus指該標本的前上頜骨牙齒及下頜最前面三顆牙齒有平行凹槽;後者的magnicornis則指較大型的鼻角。[9]:2,21這兩個物種有效性在後續研究中多次受到質疑。2000年布里特(Brooks Britt)等人表示鼻角種正模標本其實是隻少年個體,兩個較大型物種則是前者的成年狀態。[12]2003年勞赫特(Oliver Rauhut)及2008年卡拉諾(Matthew Carrano)和桑普森(Scott Sampson)評估麥德森和韋爾斯所引用的解剖學特徵,來表明這些另外物種只是個體發育(與年齡有關)或個體間形態上的差異。[13][14]:192


1992年在猶他莫爾東南方的瑪瑙盆地採石場(Agate Basin Quarry)發現了更進一步的標本BYUVP 12893,目前尚待描述。該標本來自已知最大隻的個體。包括頭骨前半部、破碎的七節骨盆處脊椎、關節相連的骨盆與薦骨。[14]:192[9]:361999年布里特記載了在懷俄明骨棺採石場發現一隻少年角鼻龍個體,由一個完整的頭骨、及其餘30%的骨骼部位(包括一個完整的骨盆)組成,其體型小於鼻角種正模的34%。[15]
除了上述五項發現外,還有來自莫里遜組第二、第四至六層帶不同地點的零碎角鼻龍遺骸的記載,[16]其中一些位在重要的化石遺址。如猶他恐龍國家紀念區的單獨右前上頜骨DNM 972;懷俄明科摩崖的大型肩胛鳥喙骨;科羅拉多旱台採石場的左肩胛鳥喙骨、脊椎和肢骨碎片;科羅拉多米蓋特摩爾採石場(Mygatt Moore Quarry)的牙齒。[9]:36
北美以外的發現
非洲
1909至1913年期間,柏林自然博物館派出德國考察隊至德屬東非(今坦尚尼亞),從敦達古魯層挖掘出了多樣化恐龍化石。[17]儘管作為非洲最重要的侏儸紀恐龍化石產地,[17]當地的大型獸腳類恐龍卻僅有少量破碎不堪的遺骸得知。[18]1920年德國古生物學家沃納·亞嫩施將從TL採石場挖到的多節背椎歸入角鼻龍未命名種(Ceratosaurus sp.)。1925年亞嫩施根據Mw採石場的破碎遺骸,包括方骨、腓骨、尾椎等碎片,命名了新種羅氏角鼻龍(Ceratosaurus roechlingi),該標本屬於一隻比鼻角種正模標本比例上更大型的個體。[18]
麥德森和韋爾斯在他們2000年的專著中證實將以上發現歸入角鼻龍。此外他們還將原本亞嫩施歸入貪食龍可能物種(Labrosaurus (?) stechowi)的牙齒改歸入角鼻龍。[9]其他研究人員質疑敦達古魯層化石的身分,指出這些標本無法辨識出任何角鼻龍的鑑定特徵。[19][20]:66[14]:192[21]2011年勞赫特發現羅氏角鼻龍及貪食龍都可能是種角鼻龍科,但缺乏可鑑定為獨立屬的鑑定特徵,因此是疑名。[21]1990年提摩西·羅威(Timothy Rowe)和賈奎斯·高錫爾提到敦達古魯25顆單獨、非常大型(長達15公分)的牙齒,據傳曾由亞嫩施提出為另一個物種碩大角鼻龍(Ceratosaurus ingens)。[19][18]然而亞嫩施其實將之歸入斑龍而非角鼻龍,也可能是單純印刷誤會。[9]:37[18]2011年勞赫特發現碩大斑龍/角鼻龍實際上與兩者都無關連,反而可能是種鯊齒龍科。[21]
歐洲
一個來自英格蘭中侏羅世地層的破碎頭骨似乎顯示出與角鼻龍相似的鼻角。1926年馮休內將頭骨描述命名成原角鼻龍,推測是角鼻龍的直系祖先。[22]現在已經知道原角鼻龍其實屬於更衍化的暴龍超科;[23]類似的鼻突結構可能是趨同演化。[14]:1852010年勞赫特等人為其建立了專屬的原角鼻龍科,且指出保存不完整的「鼻角」實際上只是更大型頭冠的最前端斷裂,一如近緣五彩冠龍的冠飾結構。[23]
2000年和2006年時,奧塔維歐·馬諦尤斯所率領的古生物學團隊描述了在葡萄牙中西部洛里尼揚層的化石ML 352,出土自洛里尼揚與托雷斯韋德拉什之間的Valmitão海岸懸崖上,包括一個右股骨、一個左脛骨、以及數個單獨的牙齒,作為角鼻龍的一個新種。[24][25]這些骨頭埋藏於黃色至棕色細粒徑砂岩,為河流氾濫沖積而成,屬於新港段(Porto Novo Member)下部層位,地質年代約啟莫里階晚期。該個體的其他骨頭SHN (JJS)-65包含左股骨、右脛骨、部分左腓骨,因懸崖持續的侵蝕作用而暴露出來。起初為私人收藏家所有,後來捐贈給托雷斯韋德拉什的自然史協會,使其成為額外的官方標本,並於2015年進行詳細描述。[26]2006年馬諦尤斯等人將該標本歸入凹齒角鼻龍。[25]2008年卡拉諾和桑普森的檢驗證實的確屬於角鼻龍,但尚不足以指定為任何特定物種。[14]:1922015年伊莉莎白·馬拉法雅(Elisabete Malafaia)等人質疑凹齒種的有效性,並將標本列在近似鼻角角鼻龍(aff. Ceratosaurus nasicornis)名下。[26]
其他記載像是瑞士穆捷的單一牙齒,起初於1920年由亞嫩施命名為梅氏貪食龍(Labrosaurus meriani),後來麥德森和韋爾斯歸入角鼻龍未命名種。[9]:35–362008年馬提亞斯·索托(Matías Soto)和丹尼爾·皮瑞(Daniel Perea)描述了烏拉圭塔瓜倫波層的牙齒(推測是前上頜骨牙冠)。其內側顯示出垂直條紋、前緣無鋸齒狀,這種特徵組合僅於角鼻龍得知,然而研究人員指出有鑑於殘骸稀缺,將歐洲及非洲材料歸入角鼻龍並不可靠,且須對其進一步謹慎評估。[27]2020年索托等人描述了更多來自同地層的角鼻龍牙齒,進而證實他們過去的解釋。[28]
特徵描述
體型

角鼻龍符合獸腳類的典型身體結構。[5]角鼻龙是一类手臂縮短、用強有力後腿移動的雙足動物。鼻角種首個發現也是正模標本的USNM 4735,不同研究估計出其體長為5.3公尺或5.69公尺。[2]:115[6]該個體是否達成熟則未能確認。[20]:661884年馬許推測這個標本體重可能是同期異特龍的一半。[3]較近期的估計則得出418公斤、524公斤、670公斤的數值。[29]後來20世紀下半所發現的另外三隻標本體型更大。第一具UMNH VP 5278麥德森曾非正式推估身長達8.8公尺,[10]後來縮減至7公尺。[30]不同研究測量的體重值為980公斤、452公斤、700公斤。[6][30][31]第二具MWC 1略小於前者,體重可能約275公斤。[31]第三具BYUVP 12893尚未經過正式描述,被稱是已發現最大型的標本,但數值並未公布。[14]:1922000年葡萄牙發現的ML 352估計身長6公尺及體重600公斤。[30]
頭骨

與身體其餘部分相比,頭骨佔了相當大的比例。[5]鼻角種正模標本的頭骨由吻尖至枕髁(寰椎附著處)測量長55公分。[2]:88該頭骨因嚴重變形使寬度難以準確復原,1920年吉爾摩版本後來被發現復原得過寬。[32]MWC 1近乎完整的頭骨測量長60公分及寬16公分;較正模標本來得加長一些。[9]:3頭骨孔洞大,使頭後部結構比其他大型獸腳類輕,但下頜更深能容納大比例的牙齒。[6]:277淚骨除了作為眶前孔(眼窩與鼻孔間的大孔)後緣也作為上緣,不同於阿貝力龍科。方骨(底部關節與下頜連接)傾斜,導致頜關節位置相對枕髁更往後移;亦造成側顳孔(眼窩後方的大孔)底部加寬。[20]:53
角鼻龍最明顯的特徵在於突出的鼻角,位於骨質鼻孔後方,沿頭骨中線處生長,由左右鼻骨癒合構成突起。[2]:82化石僅保存骨質核心部分,相信生前可能覆蓋著角質鞘。雖然角核底部平滑,上面三分之二表面布滿皺褶與多道凹槽,生前可能分布著血管。正模標本角核長13公分及寬2公分,接著大幅縮減至長7公分及寬1.2公分。[2]:82MWC 1的鼻角則較長且低矮。[9]:3生前若覆蓋角質鞘將使鼻角整體更為加長。[33]鼻角後方的鼻骨構成一橢圓凹陷;該凹陷連同鼻角均作為角鼻龍及近緣物種間的區別特徵。[14]:192除了大型鼻角,在雙眼前方有稍小、半圓形骨質脊狀物,由淚骨構成,類似異特龍。[31]年輕個體的三支角都小於成體,且角核兩邊尚未完全癒合。[15]
_-_AMNH_-_DSC06292.JPG)
構成吻尖的前上頜骨每側僅三顆牙齒,比許多獸腳類都少。[20]:52正模標本上頜骨兩側各有15顆刀刃狀牙齒,前八顆特長且厚實,第九顆起尺寸漸小。如同典型獸腳類,側緣有小鋸齒狀,正模標本每5公厘有約10個小鋸齒突。[2]:92MWC 1上頜骨僅有11至12顆牙齒、UMNH VP 5278每側則有12顆;後者牙齒更加厚實與彎曲。[9]:3,27全部標本上頜牙冠都特長;UMNH VP 5278的長達9.3公分,與下頜最矮牙齒同高;正模標本的長7公分而超越下頜最矮齒。其餘獸腳類僅銳頜龍有相似狀況。[34]相較之下某些阿貝力龍科牙冠特別短。[20]:92正模標本下頜齒骨每側有15顆牙齒,保存狀況很糟。MWC 1和UMNH VP 5278每側僅有11顆,後者顯示下頜齒相較於上頜比較直且沒那麼厚實。[9]:3,21
顱後骨骼

因為正模標本保存的脊柱存在許多段空缺,確切的脊椎數目無法確定。在薦骨之前,頸部及背部由至少20節脊椎構成。頸部中部椎體長且高,前後部椎體則長度短於高度。朝上的神經棘相對大型,背椎神經棘高度與椎體長度相當。薦骨由六個薦椎構成,朝上呈拱狀,中部椎體高度大幅縮減,在其他某些角鼻龍類也可見此情況。[20]:55–58尾部佔全身長度的一半,由大約50節尾椎構成。正模標本尾巴長2.84公尺。[3][2]:115尾部縱向很高,因為神經棘和人字骨都有加長。如同其他恐龍的尾部功能,作為平衡身體,並含有大型尾股肌,運動時負責向前的推進力,肌肉收縮時將大腿上部往後拉。[20]:55–58

肩胛骨跟鳥喙骨融合成單一塊骨頭,彼此間無任何明顯界線。[20]:58鼻角種正模保存了關節相連的左前肢,含有不完整的手部,雖然清修時造成關節脫落,但研究人员事先製作了石膏模型以記錄骨頭的原始相對位置。沒有任何標本保存下腕骨,使一些研究人員提出角鼻龍可能已經失去這項構造。2016年馬修·卡拉諾(Matthew Carrano)和喬納·休尼爾(Jonah Choiniere)根據模型呈現前臂骨與掌骨之間縫隙及其中的表面紋理來判斷,提出存在一或多個軟骨型的腕骨的可能性。[35]大多數較進化的獸腳類只有三根手指,但角鼻龍有四根手指,其中第四指明顯縮短。第一及第四掌骨短,第二掌骨略長於第三。不同於多數基礎獸腳類,掌骨尤其第一指比例特別短。正模標本只保存了第二、三、四指的第一指骨,指骨總數和爪骨未知。第一掌骨結構顯示該指生前也存在指骨。腳部由三趾承重,為第二至四趾,通常獸腳類第一趾縮短退化懸空,正模中沒有保存下來。1884年馬許假設角鼻龍遺失第一趾構造,但1920年吉爾摩指出第二蹠骨的附著處證實存在第一趾。[2]:112
角鼻龍在獸腳類中的獨特之處在於,沿身體中線生長著小型、加長、形狀不規則的皮甲。在第4、5頸椎、第4至10尾椎神經棘上方保存了皮甲,可能構成連續一排,從頭骨後方開始延伸至尾巴大部分。1920年吉爾摩推測圍岩中呈現的皮甲位置可能反映了生前狀態。尾巴上方的皮甲和神經棘間相隔25至38公厘,可能代表其間存在皮膚或肌肉,至於頸部皮甲與神經棘較靠近。除身體中線皮膚還有其他皮甲,如正模的58×70公厘大致四邊形骨板,其分布於身體的位置未知。[2]:113–114UMNH VP 5278標本也發現許多皮甲,描述為不規則狀。雖然多數皮甲發現位置與骨骼距離不超過5公尺,卻與任何脊椎無關連跡象,這和正模的情況不同,難以判斷其生前部位。[9]:32
分類系統

馬許在初始論文及後續研究中指出角鼻龍在當時已知獸腳類前所未見的特徵。[14]:185其中兩項特徵:癒合的骨盆、併蹠骨,是現代鳥類已知的特徵,根據馬許所述,這是表明恐龍與鳥類之間關係密切的證據之一。[36]為了將角鼻龍與異特龍、斑龍、虛骨龍區別開來,馬許為其建立了新的專屬分類群:角鼻龍下目/角鼻龍類與角鼻龍科,並作為底下唯一物種。[14]:1851892年馬許在化石戰爭的死對頭寇普對其觀點提出質疑,他認為鼻角等獨有特徵僅能支持作為一個新種,而無法區別成為獨立的屬,於是他將鼻角種歸入斑龍屬,成為鼻角斑龍(Megalosaurus nasicornis)。[37]

雖然後續研究皆視角鼻龍為一獨立屬,[2]:76在接下來一個世紀以來其親緣關係持續引發爭議。角鼻龍科和角鼻龍下目都未被廣泛接受,且僅有其他少數幾種所知甚少的物種被發現。多年來角鼻龍曾在不同文獻被歸入各種分類,如恐齒龍科、斑龍科、虛骨龍類、肉食龍下目、恐齒龍超科。[9]:21920年吉爾摩的重新檢驗認為角鼻龍是三疊紀之後已知最原始的獸腳類,與當時已知任何其他同時代的獸腳類都不近緣,理應分在自己專屬的角鼻龍科中。[2]:76但直到1980年代系統發生學分析的建立,馬許將角鼻龍類作為一獨特分類群的觀點才終於得到支持。1985年南美新發現的阿貝力龍和食肉牛龍被發現與角鼻龍近緣。1986年高錫爾辨識出腔骨龍超科與角鼻龍近緣,不過在近期研究該演化支通常被排除於角鼻龍類之外。自此之後,更多角鼻龍類的新物種相繼被發現。[14]:185

角鼻龍類從獸腳類演化階段的非常早期就與另一條演化至鳥類的堅尾龍類分化開來,於是常被當作相對基礎的演化支。[38]角鼻龍類的範疇底下包含了兩個更進化的分支西北阿根廷龍科和阿貝力龍科,兩者一同位於阿貝力龍超科內;另外還有一些較基礎的物種,如輕龍、三角洲奔龍、角鼻龍。角鼻龍本身在基礎角鼻龍類的確切位置至今仍有爭議。某些分析認為角鼻龍是所有基礎物種中相對最進階的,並與阿貝力龍超科互成姊妹群。[14]:187[39]2004年奧利弗·勞赫特(Oliver Rauhut)提出銳頜龍與角鼻龍構成姊妹關係,因為兩屬都以特長的上頜齒為鑑定特徵,他把兩屬歸到角鼻龍科,[34]後續研究皆採納此觀點,[40][41][42][33]如以下演化樹取自2012年迭哥波和勞赫特:[40]
| 角鼻龍類 Ceratosauria |
| |||||||||||||||||||||||||||||||||||||||||||||
2017年王碩等人認為西北阿根廷龍科並非如過去所想的處於阿貝力龍超科底下,而是比角鼻龍更為基礎。因為西北阿根廷龍科已被當作定義阿貝力龍超科及阿貝力龍科的固定依據,於是這樣的調整將使得每個演化支容納進更大範圍的分類群,其中也包含角鼻龍本身。2018年拉斐爾·德寇特(Rafael Delcourt)的後續研究支持前述論點,但指出需用從來都很少使用的角鼻龍超科來當作阿貝力龍超科的異名取代之。至於阿貝力龍科,德寇特為其指定出一種排除掉角鼻龍的新定義,而得以延用舊名。王等人還另外發現角鼻龍、銳頜龍、阿根廷的始阿貝力龍構成一個演化支。[42]德寇特以角鼻龍科代指這個範圍的演化支,並建議將角鼻龍科的定義修正為與角鼻龍接近、而與食肉牛龍較遠的所有物種。其支序圖如下所示:[33]
| 角鼻龍超科 Ceratosauroidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
古生物學
棲位與食性

在莫里遜層與洛里尼揚層,角鼻龍的化石經常與其他大型獸腳類一同發現,包括斑龍科的蠻龍、[43]異特龍科的異特龍。科羅拉多花園公園除了角鼻龍也有異特龍。科羅拉多旱台採石場及猶他克里夫蘭羅伊採石場分別發現至少三種大型獸腳類:角鼻龍、異特龍、蠻龍。[32][44]同樣在懷俄明科摩崖及鄰近地區也產出角鼻龍、異特龍、還有至少一種大型斑龍科。[45]角鼻龍是這些獸腳類動物群中數量極其稀有者,在異特龍和角鼻龍分布重疊區域,前者數量幾乎是後者的7.5倍之多。[46]
許多篇研究嘗試如何解釋這些共棲掠食者如何降低直接競爭機率。1998年唐納·韓德森(Donald Henderson)提到角鼻龍和至少兩個潛在的異特龍物種共存,稱後兩者在形態上有所分化:一種吻部縮短、頭骨較高且寬、牙齒短而後彎;另一種吻部較長、頭骨低矮、牙齒長而筆直。總體來說,若共棲物種之間的形態、生理構造、生活行為等方面的相似程度越高,彼此競爭就越激烈。韓德森結論道:短吻異特龍與長吻異特龍、角鼻龍有著不同棲位;短吻形態減少咬合產生的應力從而增加咬合力,類似貓科的狀況。至於長吻型異特龍和角鼻龍的長窄吻部及長尖牙則適合迅速猛烈的小範圍咬合,類似犬科。韓德森指出由於角鼻龍和長吻異特龍的頭骨形態十分相似,顯示兩者可能會直接競爭。根據角鼻龍在克里夫蘭羅伊採石場的罕見程度,以及當地長吻異特龍的繁盛情況來判斷,前者可能受不了後者競爭而幾乎被趕出棲地。但花園公園和旱台的角鼻龍似乎比較常見,並與短吻異特龍共存。[32]

另外韓德森認為角鼻龍也許能夠透過不一樣的獵物偏好來避免競爭;其演化出極細長的牙齒可能是與長吻異特龍競爭下的產物,在作為食腐者時兩者可能選擇取食屍體的不同部位。角鼻龍的加長牙齒也能當作物種間的視覺識別或其他社會功能。再者這些獸腳類同樣趨向體型巨大化也會降低競爭機率,因為當體型增加能選擇的獵物種類、範圍亦隨之增廣。[32]
2006年福斯特和丹尼爾·舒爾(Daniel Chure)認同韓德森的理論:有鑑於角鼻龍和異特龍共棲並捕食同類型獵物的事實,所以可能各自採取不同社食策略以避免競爭。據他們表示頭骨、牙齒、前肢在比例上有所差異也代表了證據支持。[46]但後續研究則質疑了異特龍的兩種變異形態差異。2010年肯尼思·卡彭特發現克里夫蘭羅伊採石場的短吻異特龍反映了極端程度的個體差異,而非不同物種的變異。[47]而且韓德森提出短吻形態的依據-花園公園的USNM 4734頭骨標本,實際上是將樣貌復原得太短的誤會。[48]
2004年羅伯特·巴克和蓋瑞·比爾(Gary Bir)提出角鼻龍主要特化來覓食水生獵物,如肺魚、鱷魚、海龜。統計了科摩崖及周圍50個地點發現的脫落牙齒,顯示角鼻龍和斑龍科的牙齒在水域鄰近區域(潮濕的氾濫平原、湖岸、沼澤)最常見。角鼻龍偶爾出現於內陸環境。至於異特龍科無論是內陸或近水都相當普遍。巴克和比爾據此結論道:角鼻龍和斑龍科必定在水域附近環境捕獵,而角鼻龍偶爾也會去內陸吃大型恐龍屍體。研究者還注意到角鼻龍與斑龍科都有著狹長、低矮、靈活的身體。與其他莫里遜層獸腳類相比,角鼻龍前部尾椎神經棘高聳、垂直而非後傾,加上尾椎下方的長型人字骨,顯示可能構成一條如鱷魚般的縱深形尾部,適合游泳。異特龍則相反,身形短高、粗壯結實、腿長,適應於開闊地區奔馳並追捕大型植食恐龍(如蜥腳類和劍龍類),但巴克和比爾推測在這些獵物隨季節發生短缺時,異特龍也會投機轉而覓食水生獵物。[45]但該理論受到2019年尹灿奎的質疑,他表示角鼻龍僅比莫里遜層其他獸腳類更有能力捕捉水生獵物,而非完全半水棲生活。[49]
1986年巴克的暢銷書《恐龍異說》(The Dinosaur Heresies)中認為上頜骨僅鬆散附著於周圍的頭骨上,能夠在顱內產生一定程度的活動範圍,稱為頭骨運動。同樣地,下頜可以相互移動,方骨外擴,使下頜於關節處撐開,讓動物能張大嘴巴以吞下大型食物。[50]2008年凱西·郝立蝶(Casey Holliday)和羅倫斯·灰墨(Lawrence Witmer)重新審視這項關於其他恐龍的假說,認為沒有任何證據支持恐龍存在肌肉驅動的頭骨運動,甚至多數恐龍都沒辦法做到。[51]

在一隻異特龍的恥骨腳(恥骨末端擴張處)發現某種別的獸腳類齒痕,可能由角鼻龍或蠻龍造成。因為該骨頭位於自身難以搆到的身體底緣並是整個身體最大塊骨頭,代表是這隻異特龍屍體被啃食。[52]麥傑特摩爾(Mygatt-Moore)採石場保存了異常高頻率的獸腳類咬痕,其中大部分來自異特龍和角鼻龍,也有更大的蠻龍或食蜥王龍。雖然植食動物屍體上的咬痕位置與被殺時或剛開始吃的位置一致,但異特龍身上也找到了其他異特龍或獸腳類留下的啃屍痕跡。與其他化石群相比之下,當地異常高密度的獸腳類咬痕,可能代表著旱季來臨掠食者們爭相搶奪緊縮的食物來源,或僅歸咎於其他化石點的採樣偏差。[53]
鼻角與皮甲功能
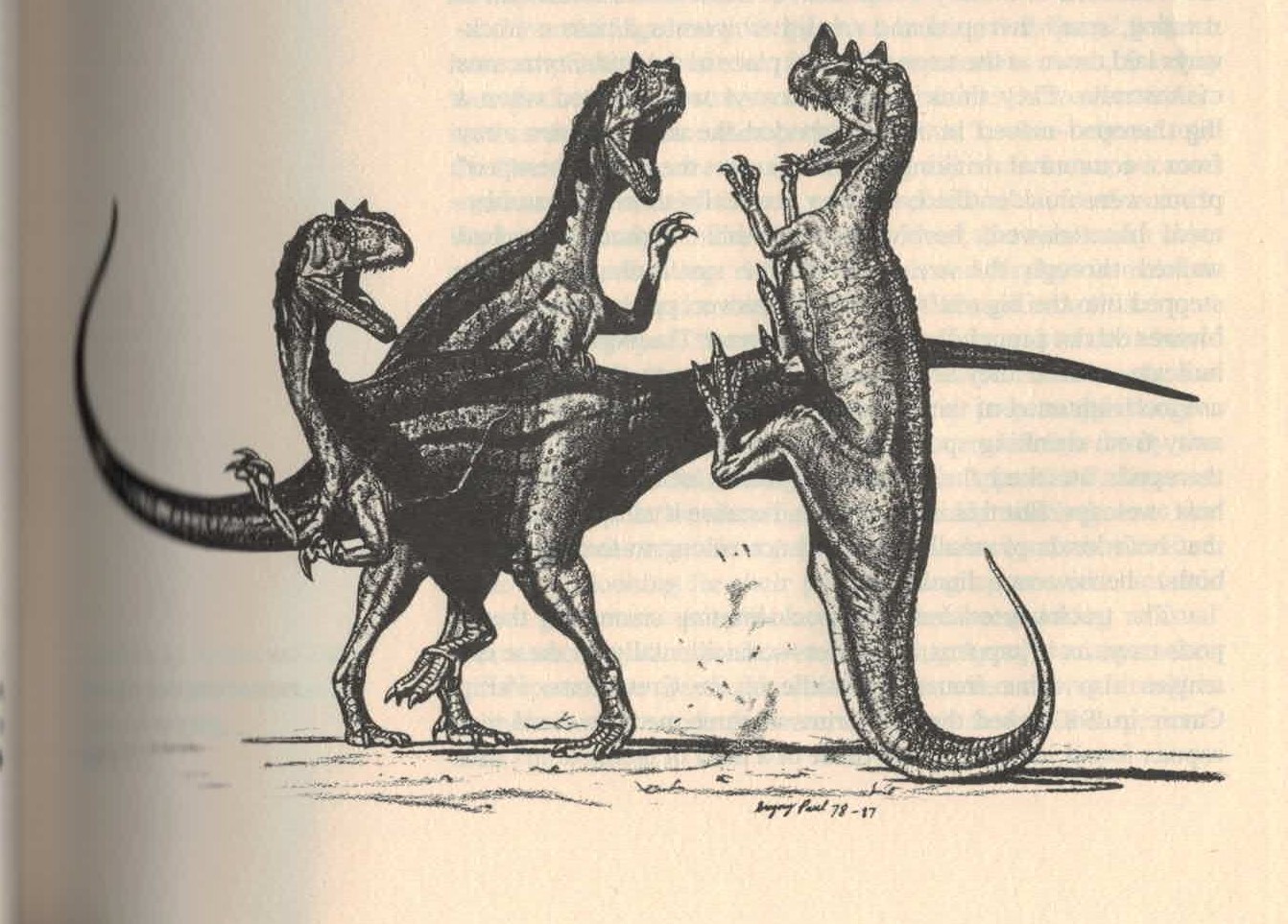
1884年馬許認為角鼻龍的鼻角是作為攻擊與防禦的強大武器,1920年吉爾摩認同其論點。[3]:331[2]:82然而現代普遍認為鼻角不太可能當作武器。[10]1985年大衛·諾曼認為鼻角可能並非用來抵禦其他掠食者,而是爭奪配偶的雄性角鼻龍種內打鬥使用。[54]1988年奎格里·保羅提出類似功能,並繪製兩隻角鼻龍進行一場非致命的互撞競賽。[6]1990年羅威和高席爾提出更進一步的論點,認為鼻角僅具有視覺展示功能,完全無法打鬥。[19]為了達到炫耀目的,鼻角可能具有鮮豔色彩。[31]另外沿身體中線生長的成排皮甲也可能有展示功能。[19]
前肢功能

角鼻龍掌骨及指骨大幅縮短,產生疑問關於手部是否仍具其他基礎獸腳類推測的抓握功能。角鼻龍類的阿貝力龍科、泥潭龍可觀察到更極端的前肢萎縮,完全失去原來的手部功能。[55]2016年卡拉諾和喬納·休尼爾(Jonah Choiniere)的角鼻龍手部解剖學研究中,指出其仍與其他基礎獸腳類形態大致雷同,就算縮短了仍保有基礎的抓握能力。雖然只有第一指骨保存下來,但根據上面的發達關節面所示,第二指骨可以靈活移動,甚至可能手指也能做到像其他基礎獸腳類那樣程度的運動。如同其他獸腳類(不含阿貝力龍科),第一指彎曲時會稍微內旋。[35]
腦部與感官

馬許可能在清修正模化石的過程中監督了腦腔模型的製作,從而得出腦部尺寸中等,但相對植食動物的要大得多。但後來頭骨被黏合在一塊,使後續研究無法檢驗該模型的準確程度。[3][2]:93
果城的MWC 1標本保存了第二件較良好的腦殼,由山德斯(Kent Sanders)和史密斯(David Smith)進行CT掃描以重建內耳結構。2005年研究人員表示角鼻龍有著一顆典型的基礎獸腳類腦腔,近似於異特龍。參與嗅覺的嗅球印痕保存良好,雖然類似異特龍,但小於暴龍;後者認為有相當靈敏的嗅覺。參與平衡感的半規管能藉以推測習慣性的頭部姿態,類似其他獸腳類;通常獸腳類中的情況雷同,顯示不同物種彼此有著差不多的運動功能。半規管最前方擴張,常見於二足動物。半規管外側的方向代表自然狀態下頭頸維持水平向。[56]
併蹠骨與古病理學
鼻角種正模的左邊第二至第四蹠骨癒合。1884年馬許寫了一篇短文探討這項當時在恐龍前所未見的特徵,並提到跟現代鳥類非常相似。[36]1890年關於角鼻龍該特徵的存在引發爭議,喬格·鮑爾推測這是骨折癒合造成的。1892年寇普認同前述論點,還認為各屬間缺乏解剖學差異,應將鼻角種視為斑龍的一種。[37]但自此後已在其他恐龍(包括比角鼻龍更基礎的物種)也觀察到非病理因素的併蹠骨。[57]1920年奧斯本說明就算併蹠骨不尋常,卻沒發現任何明顯的骨骼異常發育,可能與病理無關。[2]:1121976年拉克維奇(Ronald Ratkevich)表示蹠骨癒合將限制動物奔跑能力,1988年保羅則反駁,指出現今許多移動迅速的動物(如地棲鳥類和有蹄類)也存在併蹠骨。[6]1999年坦克(Darren Tanke)和羅斯柴爾德(Bruce Rothschild)進行分析發現該併蹠骨的確為病理因素,證實早先鮑爾的論點。[57]其他與角鼻龍有關的病理報告還有腳部的疲勞性骨折、[58]未知種的一顆斷牙呈現持續磨耗跡象。[57]
古生態學與古地理學
年代與環境

所有北美的角鼻龍都出土於莫里遜層,這是美西一系列由淺海和沖積層組成的沉積岩,也是大陸上恐龍化石最繁盛的產地。根據放射性定年法測量該岩層年代範圍,從最底層的1億5630萬年前[59]到最頂層的1億4680萬年前[60],橫跨晚牛津階、啟莫里階、到早提通階。已知的角鼻龍來自啟莫里階和提通階層帶。[20]:49莫里遜層的環境為乾濕季分明的半乾旱氣候。莫里遜盆地一路由新墨西哥州延伸到亞伯達省和薩克齊萬省,是洛磯山脈前山開始西推時形成。東向流域盆地的沉積物由溪流、河川所搬運,沉積成低地沼澤、湖泊、河道、氾濫平原。[61]莫里遜層的年代與葡萄牙洛里尼揚層、坦尚尼亞敦達古魯層大致相近。[62]
北美
莫里遜層保存了由巨型蜥腳類佔據優勢的環境與時代記錄。[63]與角鼻龍共存的恐龍包括:獸腳類剖齒龍、史托龍、長臂獵龍、虛骨龍、馬許龍、嗜鳥龍、異特龍、食蜥王龍、蠻龍;蜥腳類迷惑龍、雷龍、春雷龍、載域龍、簡棘龍、腕龍、圓頂龍、梁龍、小梁龍、重龍、超龍、極巨龍;裝甲類劍龍、西龍、阿爾科瓦龍、怪嘴龍、邁摩爾甲龍;鳥腳類侏儒龍、橡樹龍、彎龍。[64]異特龍標本數量佔莫里遜層所有獸腳類的70至75%,處於當地食物鏈頂端。[44]其他共享棲地的脊椎動物還有輻鰭魚、青蛙、蠑螈、烏龜的多塞龜、喙頭目、蜥蜴、陸生和水生鱷形超目如跳鱷、數種翼龍類包括抓頜龍和買薩翼龍。雙殼綱、水生蝸牛的殼也很常見。藻類、真菌、蘚苔、木賊、蘇鐵、銀杏、數種針葉樹化石透露了當時的植相狀況。植被景觀包括樹蕨等蕨類構成的濱岸林及充斥著南洋杉屬之類的稀樹莽原。[65]
歐洲
葡萄牙洛里尼揚層新港段(Porto Novo Member)發現角鼻龍的破碎遺骸透露了該物種也許存在於此地。洛里尼揚層的恐龍生態組成結構與莫里遜層幾乎一樣,或至少都是類似物種。[62]除了角鼻龍,葡萄牙岩層也同樣出土過來自北美的異特龍和蠻龍,至於洛里尼揚諾龍只在葡萄牙發現過。新港段的植食性恐龍包括蜥腳類的迪涅魯龍、洛里尼揚龍、茲必龍;劍龍類的米拉加亞龍。[66][24][25]在晚侏羅世,歐洲與北美之間僅隔著剛形成的狹窄大西洋,而葡萄牙所在的伊比利半島當時是一座大型孤島,與歐洲其他地區隔離開來。根據馬諦尤斯等人所述,葡萄牙與北美獸腳類種群的相似程度,代表之間曾經存在過陸橋使動物得以互相遷徙。[24][25]不過馬拉法雅等人提出不同、更為複雜的情境假設,因為葡萄牙其他動物如蜥腳類、海龜、鱷類都與北美有明顯差異,代表當陸塊開始分開時,某些物種是透過交換遷徙來的,某些則在當地產生特有化。[26]
文化意義
公眾形象
角鼻龍是知名的恐龍之一。因為來自美國本土,又早在19世紀就發表,足以長期發展成為科普媒體中代表性的經典史前物種。史密松寧學會博物館的展覽品提供了加持,因為當時那邊的恐龍廳展品很少,而角鼻龍獨特的鼻角和背脊賦予了一種迷人的「龍狀」外表令人印象深刻。[67]
繪畫
.jpg)


1899年邦德(Frank Bond)在威柏·克林頓·奈特指導下繪製了可能是歷史上首個角鼻龍復原圖,但直到1920年才透過吉爾摩公開。[2]1901年費德里克·奧古斯特斯·盧卡斯出版一本關於滅絕動物的科普書《Animals of the Past》,當中有著名童書畫喬瑟夫·格里森的復原圖,[68]他在查爾斯·R·奈特監督下繪製。吉爾摩曾表示這是最為成功的復原,還將圖放在博物館骨架旁展示。[2]1911年亨利·內維爾·哈欽森的《Extinct Monsters and Creatures of Other Days: a popular account of some of the larger forms of ancient animal life》出現由喬瑟夫·斯密繪的角鼻龍插畫,顯然參考了馬許的錯誤復原。[69]1915年吉爾摩製作了角鼻龍的黏土模型。[70]
1920年代以後,角鼻龍形象長期固化,後續的畫家皆參考格里森或斯密,逐漸完全脫離動物的真實形象。直到1980年代突然起了變革,奎格里·保羅觀察了正模標本,將角鼻龍詮釋為行動活躍的水平姿態,更貼近現代形象。其中一幅 (页面存档备份,存于互联网档案馆)角鼻龍以尾巴撐地抬起雙腳踢擊兩隻異特龍,類似袋鼠的行為。另一幅 (页面存档备份,存于互联网档案馆)依照他對鼻角功能的論點,展現了角鼻龍透過鼻角側擊對抗同類。這些成為新式古生物形象的代表性作品,並在1990年代佔據主流地位。[6]
{kind=link}
{kind=link}
{kind=link}
{kind=link}
媒體作品

角鼻龍早期在文學作品的亮相,可追溯到1918年愛德加·萊斯·巴勒斯的科幻小說《時光遺忘之境》(The Land That Time Forgot)。[71]1942年迪諾·布扎蒂所創作的美文中篇小說《屠龍記》(l'Uccisione del drago)中描述了最後倖存的角鼻龍。[72]角鼻龍亦成為某些神秘生物學的題材,在1908年喬吉斯·德派的作品《鷓鴣溪怪獸》(The Monster of Partridge Creek)描繪育空地區出沒的大型二足怪物形象即參考了角鼻龍;[73][74]但後世批判了恐龍生活在極圈的可能性。[75]
20世紀角鼻龍開始經常出現於大螢幕,使肉食恐龍的代表形象廣泛為人所知。1914年D·W·格里菲斯的電影《原始人》(Brute Force)中角鼻龍作為威脅穴居部落的史前動物之一,該作因使用機械模型而被視為史上第一部有實體恐龍登場的電影。[76]1940年迪士尼的《幻想曲》(Fantasia)中的《春之祭》片段 (页面存档备份,存于互联网档案馆),角鼻龍攻擊陷入泥淖的蜥腳類和劍龍。1948年的電影《未知島 (页面存档备份,存于互联网档案馆)》也出現角鼻龍。1966年的電影《史前百萬年 (页面存档备份,存于互联网档案馆)》有角鼻龍跟三角龍的打鬥場景,但這兩種實際上不可能共存;片中的恐龍由雷·哈利豪森的定格動畫所製作。1975年改編同名小說的電影《時光遺忘之境》《The Land That Time Forgot》及1977年的續集《時光遺忘之人》也出現了角鼻龍。2001年的《侏羅紀公園3》(Jurassic Park III)一隻紅色的角鼻龍於主角們在棘龍糞便尋找電話的片段中短暫登場。同系列動畫《白堊紀冒險營》(Camp Cretaceous)角鼻龍再度回歸。[77]角鼻龍亦在幾部科普作品有亮相。1956年的紀錄片《動物世界》(The Animal World)以定格動畫展現角鼻龍殺死劍龍,後同類爭食雙雙跌下懸崖被沖走。[78]2001年探索頻道的紀錄片《恐龍紀元》(When Dinosaurs Roamed America)的晚侏羅世段落,角鼻龍先登場追捕橡樹龍並殺死一隻,然後試圖襲擊劍龍被打倒而放棄,最後再度追捕橡樹龍時被異特龍偷襲咬死。[79]
參考來源
- ^ Brinkman, P.D. The Second Jurassic Dinosaur Rush. Museums and Paleontology in America at the Turn of the Twentieth Century. University of Chicago Press. 2010: 10. ISBN 978-0-226-07472-6.
- ^ 2.00 2.01 2.02 2.03 2.04 2.05 2.06 2.07 2.08 2.09 2.10 2.11 2.12 2.13 2.14 2.15 2.16 2.17 2.18 2.19 2.20 2.21 2.22 Gilmore, C.W. Osteology of the carnivorous Dinosauria in the United States National Museum, with special reference to the genera Antrodemus (Allosaurus) and Ceratosaurus (PDF). Bulletin of the United States National Museum. 1920, 110 (110): 1–154 [2022-06-20]. doi:10.5479/si.03629236.110.i. hdl:2027/uiug.30112032536010
 . (原始内容存档 (PDF)于2021-11-09).
. (原始内容存档 (PDF)于2021-11-09).
- ^ 3.0 3.1 3.2 3.3 3.4 Marsh, O.C. Principal characters of American Jurassic dinosaurs, part VIII: The order Theropoda (PDF). American Journal of Science. 1884, 27 (160): 329–340 [2022-06-20]. Bibcode:1884AmJS...27..329M. doi:10.2475/ajs.s3-27.160.329. (原始内容存档 (PDF)于2023-02-15).
- ^ Simpson, D.P. Cassell's Latin Dictionary 5. London: Cassell Ltd. 1979: 153, 387 [1854]. ISBN 978-0-304-52257-6.
- ^ 5.0 5.1 5.2 Marsh, O.C. Restorations of Claosaurus and Ceratosaurus. American Journal of Science. 1892, 44 (262): 343–349 [2022-06-20]. Bibcode:1892AmJS...44..343M. doi:10.2475/ajs.s3-44.262.343. hdl:2027/hvd.32044107356040 . (原始内容存档于2018-07-22).
- ^ 6.0 6.1 6.2 6.3 6.4 6.5 6.6 Paul, Gregory S. Ceratosaurs. Predatory Dinosaurs of the World. Simon & Schuster. 1988: 274–279. ISBN 978-0-671-61946-6.
- ^ Jacqueline, T. David Koch Donates $35 Million to National Museum of Natural History for Dinosaur Hall. The Washington Post. May 3, 2012 [December 4, 2017]. (原始内容
 存档于2017-07-19).
存档于2017-07-19).
- ^ 8.0 8.1 The Good, Bad and Ugly: Dismantling Historic Fossil Displays, Part Two. Digging the Fossil Record: Paleobiology at the Smithsonian. Smithsonian National Museum of Natural History. [December 4, 2017]. (原始内容存档于October 10, 2017).
- ^ 9.00 9.01 9.02 9.03 9.04 9.05 9.06 9.07 9.08 9.09 9.10 9.11 9.12 9.13 9.14 Madsen, J.H.; Welles, S.P. Ceratosaurus (Dinosauria, Theropoda): A Revised Osteology. Utah Geological Survey. 2000: 1–80. ISBN 978-1-55791-380-7.
- ^ 10.0 10.1 10.2 Glut, D.F. Ceratosaurus. Dinosaurs: The Encyclopedia. McFarland & Company. 1997: 266–270. ISBN 978-0-89950-917-4.
- ^ Tidwell, V. Restoring crushed Jurassic dinosaur skulls for display. The Continental Jurassic. Museum of Northern Arizona Bulletin. 1996, 60.
- ^ Britt, B.B.; Chure, D.J.; Holtz, T.R., Jr.; Miles, C.A.; Stadtman, K.L. A reanalysis of the phylogenetic affinities of Ceratosaurus (Theropoda, Dinosauria) based on new specimens from Utah, Colorado, and Wyoming. Journal of Vertebrate Paleontology. 2000, 20 (suppl): 32A. doi:10.1080/02724634.2000.10010765.
- ^ Rauhut, O.W.M. The interrelationships and evolution of basal theropod dinosaurs. Special Papers in Palaeontology. 2003: 25.
- ^ 14.00 14.01 14.02 14.03 14.04 14.05 14.06 14.07 14.08 14.09 14.10 Carrano, M.T.; Sampson, S.D. The Phylogeny of Ceratosauria (Dinosauria: Theropoda). Journal of Systematic Palaeontology. 2008, 6 (2): 183–236. ISSN 1477-2019. doi:10.1017/S1477201907002246.
- ^ 15.0 15.1 Britt, B.B.; Miles, C.A.; Cloward, K.C.; Madsen, J.H. A juvenile Ceratosaurus (Theropoda, Dinosauria) from Bone Cabin Quarry West (Upper Jurassic, Morrison Formation), Wyoming. Journal of Vertebrate Paleontology. 1999, 19 (Supplement to No 3): 33A. doi:10.1080/02724634.1999.10011202.
- ^ Foster, J. Appendix. Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. 2007: 327–329. ISBN 978-0-253-34870-8.
- ^ 17.0 17.1 Zils, C.W.; Moritz, A.; Saanane, C. Tendaguru, the most famous dinosaur locality of Africa. Review, survey and future prospects. Documenta Naturae. 1995, 97: 1–41.
- ^ 18.0 18.1 18.2 18.3 Janensch, W. Die Coelurosaurier und Theropoden der Tendaguru-Schichten Deutsch-Ostafrikas. Palaeontographica. 1925,. Supplement VIII: 1–100 (德语).
- ^ 19.0 19.1 19.2 19.3 Rowe, T.; Gauthier, J. Ceratosauria. Weishampel, D.B.; Dodson, P.; Osmólska, H. (编). The Dinosauria. University of California Press. 1990: 151–168. ISBN 978-0-520-06726-4.
- ^ 20.0 20.1 20.2 20.3 20.4 20.5 20.6 20.7 20.8 Tykoski, R.S.; Rowe, T. Ceratosauria. Weishampel, D.B.; Dodson, P.; Osmólska, H. (编). The Dinosauria: Second Edition. University of California Press. 2004: 47–70. ISBN 978-0-520-24209-8.
- ^ 21.0 21.1 21.2 Rauhut, O.W.M. Theropod dinosaurs from the Late Jurassic of Tendaguru (Tanzania). Special Papers in Palaeontology. 2011, 86: 195–239.
- ^ Huene, F.v. On several known and unknown reptiles of the order Saurischia from England and France. Annals and Magazine of Natural History. 1926,. Serie 9 (17): 473–489. doi:10.1080/00222932608633437.
- ^ 23.0 23.1 Rauhut, O.W.M.; Milner, A.C.; Moore-Fay, S. Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England. Zoological Journal of the Linnean Society. 2010, 158 (1): 155–195. doi:10.1111/j.1096-3642.2009.00591.x .
- ^ 24.0 24.1 24.2 Mateus, O.; Antunes, M.T. Ceratosaurus (Dinosauria: Theropoda) in the Late Jurassic of Portugal. 31st International Geological Congress, Abstract Volume (Rio de Janeiro). 2000.
- ^ 25.0 25.1 25.2 25.3 Mateus, O.; Walen, A.; Antunes, M.T. The large theropod fauna of the Lourinhã Formation (Portugal) and its similarity to the Morrison Formation, with a description of a new species of Allosaurus. Foster, J.R.; Lucas, S.G. (编). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science, Bulletin 36. 2006.
- ^ 26.0 26.1 26.2 Malafaia, E.; Ortega, F.; Escaso, F.; Silva, B. New evidence of Ceratosaurus (Dinosauria: Theropoda) from the Late Jurassic of the Lusitanian Basin, Portugal. Historical Biology. October 3, 2015, 27 (7): 938–946. ISSN 0891-2963. doi:10.1080/08912963.2014.915820.
- ^ Soto, M.; Perea, D. A ceratosaurid (Dinosauria, Theropoda) from the Late Jurassic–Early Cretaceous of Uruguay. Journal of Vertebrate Paleontology. 2008, 28 (2): 439–444. doi:10.1671/0272-4634(2008)28[439:acdtft]2.0.co;2.
- ^ Soto, Matías; Toriño, Pablo; Perea, Daniel. Ceratosaurus (Theropoda, Ceratosauria) teeth from the Tacuarembó Formation (Late Jurassic, Uruguay). Journal of South American Earth Sciences. November 1, 2020, 103: 102781. Bibcode:2020JSAES.10302781S. ISSN 0895-9811. S2CID 224842133. doi:10.1016/j.jsames.2020.102781 (英语).
- ^ Therrien, F.; Henderson, D.M. My theropod is bigger than yours ... or not: estimating body size from skull length in theropods. Journal of Vertebrate Paleontology. 2007, 27 (1): 108–115. doi:10.1671/0272-4634(2007)27[108:mtibty]2.0.co;2.
- ^ 30.0 30.1 30.2 Paul, Gregory S. The Princeton Field Guide to Dinosaurs 2nd. Princeton University Press. 2016: 84. ISBN 978-0-691-16766-4.
- ^ 31.0 31.1 31.2 31.3 Foster, J. Gargantuan to Minuscule: The Morrison Menagerie, Part II. Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. 2007: 162–242. ISBN 978-0-253-34870-8.
- ^ 32.0 32.1 32.2 32.3 Henderson, D.M. Skull and tooth morphology as indicators of niche partitioning in sympatric Morrison Formation theropods. Gaia. 1998, (15): 219–226.
- ^ 33.0 33.1 33.2 Delcourt, Rafael. Ceratosaur palaeobiology: new insights on evolution and ecology of the southern rulers. Scientific Reports. 2018, 8 (1): 9730. Bibcode:2018NatSR...8.9730D. PMC 6021374 . PMID 29950661. doi:10.1038/s41598-018-28154-x.
- ^ 34.0 34.1 Rauhut, O.W.M. Provenance and anatomy of Genyodectes serus, a large-toothed ceratosaur (Dinosauria: Theropoda) from Patagonia. Journal of Vertebrate Paleontology. 2004, 24 (4): 894–902. doi:10.1671/0272-4634(2004)024[0894:paaogs]2.0.co;2.
- ^ 35.0 35.1 Carrano, M.T.; Choiniere, J. New information on the forearm and manus of Ceratosaurus nasicornis Marsh, 1884 (Dinosauria, Theropoda), with implications for theropod forelimb evolution. Journal of Vertebrate Paleontology. 2016, 36 (2): –1054497. doi:10.1080/02724634.2015.1054497.
- ^ 36.0 36.1 Marsh, O.C. On the united metatarsal bones of Ceratosaurus. American Journal of Science. 1884, 28 (164): 161–162 [2022-06-20]. Bibcode:1884AmJS...28..161M. doi:10.2475/ajs.s3-28.164.161. (原始内容存档于2020-06-24).
- ^ 37.0 37.1 Cope, E.D. On the Skull of the Dinosaurian Lælaps incrassatus Cope. Proceedings of the American Philosophical Society. 1892, 30 (138): 240–245. ISSN 0003-049X. JSTOR 983173.
- ^ Hendrickx, C.; Hartman, S.A.; Mateus, O. An overview of non-avian theropod discoveries and classification. PalArch's Journal of Vertebrate Palaeontology. 2015, 12 (1): 1–73.
- ^ Xu, X.; Clark, J.M.; Mo, J.; Choiniere, J.; Forster, C.A.; Erickson, G.M.; Hone, D.W.; Sullivan, C.; Eberth, D.A.; Nesbitt, S.; Zhao, Q. A Jurassic ceratosaur from China helps clarify avian digital homologies (PDF). Nature. 2009, 459 (7249): 940–944 [2022-06-20]. Bibcode:2009Natur.459..940X. PMID 19536256. doi:10.1038/nature08124. (原始内容存档 (PDF)于2017-09-22).
- ^ 40.0 40.1 Pol, D.; Rauhut, O.W.M. A Middle Jurassic abelisaurid from Patagonia and the early diversification of theropod dinosaurs. Proceedings of the Royal Society B: Biological Sciences. 2012, 279 (1804): 3170–5. PMC 3385738 . PMID 22628475. doi:10.1098/rspb.2012.0660.
- ^ Delcourt, R. Revised morphology of Pycnonemosaurus nevesi Kellner & Campos, 2002 (Theropoda: Abelisauridae) and its phylogenetic relationships. Zootaxa. 2017, 4276 (1): 1–45. PMID 28610214. doi:10.11646/zootaxa.4276.1.1.
- ^ 42.0 42.1 Wang, S.; Stiegler, J.; Amiot, R.; Wang, X.; Du, G.-H.; Clark, J.M.; Xu, X. Extreme ontogenetic changes in a ceratosaurian theropod (PDF). Current Biology. 2017, 27 (1): 144–148. PMID 28017609. doi:10.1016/j.cub.2016.10.043 .
- ^ Hendrickx, Christophe; Mateus, Octávio. Torvosaurus gurneyi n. sp., the Largest Terrestrial Predator from Europe, and a Proposed Terminology of the Maxilla Anatomy in Nonavian Theropods. PLOS ONE. March 5, 2014, 9 (3): e88905. Bibcode:2014PLoSO...988905H. ISSN 1932-6203. PMC 3943790 . PMID 24598585. doi:10.1371/journal.pone.0088905 (英语).
- ^ 44.0 44.1 Foster, J.R. Paleoecological analysis of the vertebrate fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A.. New Mexico Museum of Natural History and Science Bulletin 23. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. 2003: 29.
- ^ 45.0 45.1 Bakker, Robert T.; Bir, Gary. Dinosaur crime scene investigations: theropod behavior at Como Bluff, Wyoming, and the evolution of birdness. Currie, P.J.; Koppelhus, E.B.; Shugar, M.A.; Wright, J.L. (编). Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Bloomington and Indianapolis: Indiana University Press. 2004: 301–342. ISBN 978-0-253-34373-4.
- ^ 46.0 46.1 Foster, J.R.; Chure, D.J. Hindlimb allometry in the Late Jurassic theropod dinosaur Allosaurus, with comments on its abundance and distribution. New Mexico Museum of Natural History and Science Bulletin. 2006, 36: 119–122.
- ^ Carpenter, Kenneth. Variation in a population of Theropoda (Dinosauria): Allosaurus from the Cleveland-Lloyd Quarry (Upper Jurassic), Utah, USA. Paleontological Research. 2010, 14 (4): 250–259. doi:10.2517/1342-8144-14.4.250.
- ^ Carpenter, Kenneth; Paul, Gregory S. Comment on Allosaurus Marsh, 1877 (Dinosauria, Theropoda): proposed conservation of usage by designation of a neotype for its type species Allosaurus fragilis Marsh, 1877. Bulletin of Zoological Nomenclature. 2015, 72: 1.
- ^ Changyu Yun. Comments on the ecology of Jurassic theropod dinosaur Ceratosaurus (Dinosauria: Theropoda) with critical reevaluation for supposed semiaquatic lifestyle (PDF). Volumina Jurassica. 2019, XVII: 111–116 [2022-07-02]. (原始内容 (PDF)存档于2022-07-12).
- ^ Bakker, R.T. The Dinosaur Heresies 2. William Morrow and Company. 1986: 523. Bibcode:1987Palai...2..523G. ISBN 978-0-688-04287-5. JSTOR 3514623. doi:10.2307/3514623.
|journal=被忽略 (帮助);|issue=被忽略 (帮助) - ^ Holliday, C.M.; Witmer, L.M. Cranial kinesis in dinosaurs: intracranial joints, protractor muscles, and their significance for cranial evolution and function in diapsids. Journal of Vertebrate Paleontology. December 12, 2008, 28 (4): 1073–1088. doi:10.1671/0272-4634-28.4.1073.
- ^ Chure, Daniel J.; Fiorillo, Anthony R.; Jacobsen, Aase. Prey bone utilization by predatory dinosaurs in the Late Jurassic of North America, with comments on prey bone use by dinosaurs throughout the Mesozoic. Gaia. 1998, 15: 227–232 [2022-07-02]. (原始内容存档于2021-11-09).
- ^ Drumheller, Stephanie K.; McHugh, Julia B.; Kane, Miriam; Riedel, Anja; D’Amore, Domenic C. High frequencies of theropod bite marks provide evidence for feeding, scavenging, and possible cannibalism in a stressed Late Jurassic ecosystem. PLOS ONE. May 27, 2020, 15 (5): e0233115. Bibcode:2020PLoSO..1533115D. ISSN 1932-6203. PMC 7252595 . PMID 32459808. doi:10.1371/journal.pone.0233115 (英语).
- ^ Norman, D.B. Carnosaurs. The Illustrated Encyclopedia of Dinosaurs. Salamander Books Ltd. 1985: 62–67. ISBN 978-0-517-46890-6.
- ^ Senter, P. Vestigial skeletal structures in dinosaurs. Journal of Zoology. 2010, 280 (4): 60–71. doi:10.1111/j.1469-7998.2009.00640.x.
- ^ Sanders, R.K.; Smith, D.K. The endocranium of the theropod dinosaur Ceratosaurus studied with computer tomography. Acta Palaeontologica Polonica. 2005, 50 (3).
- ^ 57.0 57.1 57.2 Molnar, R.E. Theropod paleopathology: a literature survey. Tanke, D.H.; Carpenter, K. (编). Mesozoic Vertebrate Life. Indiana University Press. 2001: 337–363.
- ^ Rothschild, B.; Tanke, D.H.; Ford, T.L. Theropod stress fractures and tendon avulsions as a clue to activity. Tanke, D.H.; Carpenter, K. (编). Mesozoic Vertebrate Life. Indiana University Press. 2001: 331–336.
- ^ Trujillo, K.C.; Chamberlain, K.R.; Strickland, A. Oxfordian U/Pb ages from SHRIMP analysis for the Upper Jurassic Morrison Formation of southeastern Wyoming with implications for biostratigraphic correlations. Geological Society of America Abstracts with Programs. 2006, 38 (6): 7.
- ^ Bilbey, S.A. Cleveland-Lloyd Dinosaur Quarry – age, stratigraphy and depositional environments. Carpenter, K.; Chure, D.; Kirkland, J.I. (编). The Morrison Formation: An Interdisciplinary Study. Modern Geology 22. Taylor and Francis Group. 1998: 87–120. ISSN 0026-7775.
- ^ Russell, D.A. An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press. 1989: 64–70. ISBN 978-1-55971-038-1.
- ^ 62.0 62.1 Mateus, O. Jurassic dinosaurs from the Morrison Formation (USA), the Lourinhã and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): A comparison. Foster, J.R.; Lucas, S.G. (编). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. 2006: 223–231.
- ^ Foster, J. (2007). "Appendix." Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. pp. 327–329.
- ^ Chure, D.J.; Litwin, R.; Hasiotis, S.T.; Evanoff, E.; Carpenter, K. The fauna and flora of the Morrison Formation: 2006. Foster, J.R.; Lucas, S.G. (编). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. 2006: 233–248.
- ^ Carpenter, K. Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus. Foster, J.R.; Lucas, S.G. (编). Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. 2006: 131–138.
- ^ Mateus, O.; Mannion, P.D.; Upchurch, P. Zby atlanticus, a new turiasaurian sauropod (Dinosauria, Eusauropoda) from the Late Jurassic of Portugal. Journal of Vertebrate Paleontology. 2014, 34 (3): 618–634. doi:10.1080/02724634.2013.822875.
- ^ A Smithsonian Dino-Celebrity Finally Tells All. [2022-07-10]. (原始内容存档于2022-07-13).
- ^ Lucas, F. A. (1901). Animals of the Past. McClure, Phillips.
- ^ Hutchinson, H. N. (1910). Extinct Monsters and Creatures of Other Days: A Popular Account of Some of the Larger Forms of Ancient Animal Life. Chapman & Hall.
- ^ The Gilmore Models: Where are they now?. [2022-07-10]. (原始内容存档于2022-07-14).
- ^ Title: The Land That Time Forgot. isfdb.org. [2022-08-01]. (原始内容存档于2022-11-20).
- ^ Giovanni Drogo (D. Buzzati), The killing of the dragon , in Oggi, Rizzoli, 3 June 1939, p. 8.
- ^ Dupuy, Georges. Newnes, George , 编. The Monster of 'Partridge Creek'. The Strand Magazine. Vol. 36 no. 211 (George Newnes Ltd.). July 1908: 73–79 –通过Internet Archive.
- ^ Dupuy, Georges. Le Monstre de 'Partridge Creek' [The Monster of "Partridge Creek"]. Je sais tout. Vol. 4 no. 39 (Paris, France: Pierre Lafitte & Cie). 15 April 1908: 403–409 [2022-08-01]. (原始内容存档于2022-09-03) –通过Bibliothèque nationale de France (法语).
- ^ Lydekker, Richard. The Monster of Partridge Creek. Zoology. Knowledge & Illustrated Scientific News (London, England). September 1908, 5 (9): 219 –通过Hathi Trust.
- ^ Primitive Man, (a.k.a. Brute Force) at silentera.com. [2022-08-01]. (原始内容存档于2020-10-28).
- ^ Ceratosaurus | Jurassic Park Wiki - Fandom. [2022-08-01]. (原始内容存档于2022-10-19).
- ^ The Animal World (页面存档备份,存于互联网档案馆)
- ^ When Dinosaurs Roamed America (页面存档备份,存于互联网档案馆)